INTRODUCTION
Melatonin and serotonin are the products of tryptophan metabolism, which are endogenous neurotransmitters and hormones. They are pleiotropic signaling chemicals that regulate a variety of physiological processes as well as counteract disease and environmental stresses in several species. They act as neurotransmitters, hormones, and bioregulators in insects controlling a variety of endocrines, behavioral (including social behavior), immunological, developmental, and protective activities, as well as biological cycles.
Melatonin is an ancient molecule found in alpha proteobacteria and cyanobacteria, indicating that it has been around since the beginning of life. Melatonin (N-acetyl-5-methoxytryptamine), an amino acid derivative, is an evolutionary conservative chemical. Melatonin is found in algae and higher plants [1], as well as nearly every invertebrate group and vertebrates [2].
Bacteria, Eucarya, unicellular and multicellular fungi, plants, and animals, including simple and complex vertebrates and invertebrates, all synthesize this pluripotent molecule with different effects in nature. Melatonin regulates several endocrines, immunological, neurological, metabolic, and defensive activities in addition to the circadian rhythm. Many researchers suggest that melatonin can be utilized as a health supplement because of its multiple biological properties, including antioxidant, anticancer, and antiaging benefits. Melatonin was identified for the first time in invertebrates in the compound eyes of an insect, the locust, Locusta migratoria [3] and it was detected using a radioimmunoassay, which was then confirmed using gas chromatography-mass spectrometry. Furthermore, melatonin is a highly efficient scavenger not only in vertebrates but also in insects [4] and dinoflagellates [5] of metabolic hydroxyl-radicals.
Serotonin, also an ancient molecule with pluripotent and diverse activities, is a result of consecutive metabolic transformations of L-tryptophan that occur in various species, including vertebrates and invertebrates such as worms, insects, fungi, plants, and unicellular organisms.
Several fascinating research studies have revealed serotoninŌĆÖs ubiquitous and overwhelming activities in vertebrates and invertebrates. Although the number of studies on the role of serotonin in insects is still few, these studies have already demonstrated the critical involvement of serotonin signaling in nearly all key physiological processes in insects. The serotonin transporter (SERT) from the fruit fly Drosophila melanogaster was identified as the first insect amine transporter.
BIOSYNTHESIS OF SEROTONIN AND MELATONIN
Melatonin and serotonin have been discovered in the heads, eyes, optic lobes, and brains of various invertebrate species. Serotonin and melatonin are produced in insects via the same pathways in vertebrates. Tryptophan-5-hydroxylase-1 (T5H-1) is the rate-limiting enzyme in the biosynthesis of serotonin, which is also involved in the biosynthesis of melatonin. Melatonin is a natural hormone that helps people cope with stressors like cold, heat, UV, and pesticides. The direct link between T5H-1 and melatonin and the underlying mechanism in organisms has yet to be explored.
Biosynthesis of serotonin
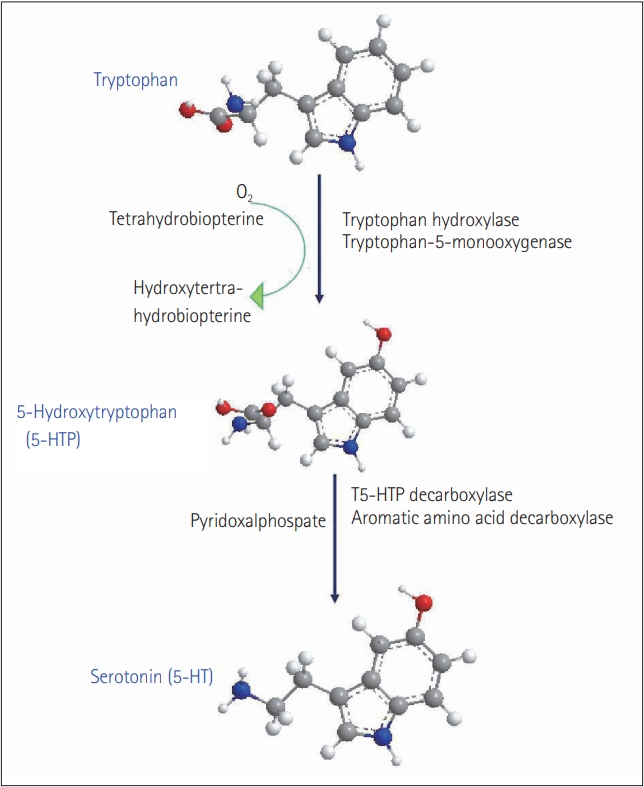
Serotonin (5-hydroxytryptamine, 5-HT) is a monoamine, implying that an aliphatic chain with two carbon oxides separates a basic amine group from an aromatic core. The important amino acid tryptophan is the first step in synthesizing 5-HT. Tryptophan hydroxylase (TPH), also known as tryptophan-5-monooxygenase, adds a hydroxyl group to tryptophan to form 5-hydroxytryptophan (5-HTP) (Figure 1) [6]. The non-heme iron and tetrahydrobiopterin-dependent aromatic amino acid hydroxylase family includes TPH, phenylalanine hydroxylase (which catalyses the hydroxylation of the aromatic side-chain of phenylalanine to produce tyrosine), and tyrosine hydroxylase (which converts tyrosine to 3,4-dihydroxyphenylalanine [DOPA]). Cofactors for these enzymes are (6R)-L-erythro-5,6,7,8-tetrahydrobiopterin and O2 [7,8].
5-HTP decarboxylase catalyzes the conversion of 5-HTP to 5-HT in the second phase of 5-HT biosynthesis (Figure 1) [9]. 5-HTP decarboxylase has been proven to be the same enzyme as dopamine decarboxylase, which catalyzes the decarboxylation stage in dopamine biosynthesis and is commonly referred to as aromatic amino acid decarboxylase (AAAD) [10].
Biosynthesis of melatonin
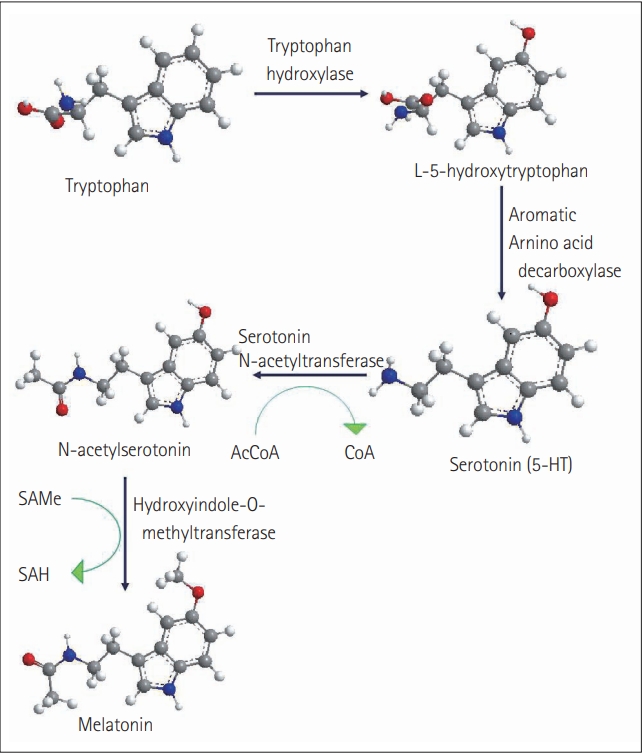
The pathways of melatonin biosynthesis in insects appear to be similar to those seen in vertebrates [2]. The amino acid precursor L-tryptophan is converted to melatonin (N-acetyl-5-methoxytryptamine) by four enzymes: TPH, AAAD, serotonin N-acetyltransferase (NAT), and hydroxyindole-O-methyltransfer (HIOMT) (Figure 2).
TPH and NAT, two enzymes involved in the multistep conversion of L-tryptophan to N-acetyl-5-ethoxytryptamine, are thought to play a critical regulatory role, with the former limiting the formation of 5-HT (serotonin) and the latter limiting the formation of melatonin [11-14]. AAAD and HIOMT appear to function primarily by mass action, while their actions may be amenable to adaptive control [13]. As an essential amino acid, L-tryptophan cannot be synthesized by vertebrate cells and must be obtained through the diet, implying dietary compositionŌĆÖs importance in the appropriate melatonin synthesis in living animals [12].
5-HTP is formed from tryptophan, decarboxylated to become 5-HT (serotonin). In the vertebrate pineal gland, 5-HT metabolism follows two primary routes: acetylation, which involves NAT activity and results in N-acetylserotonin (NAS), the direct precursor of melatonin, and oxidative deamination, which involves monoamine oxidase (MAO) activity. N-acetylation is the predominant enzymatic route at least in insects [15]. NAT is found in several species, including insects, where it is involved in the inactivation of monoamine neurotransmitters and the formation of catecholamine intermediates required for sclerotization of the insect cuticle. It is unclear what caused the phylogenetic shift from N-acetylation to MAO as an enzyme system for biogenic amine metabolism. Many insects, however, have evolved to avoid the elimination of catecholamine metabolites in aqueous excreta. Water conservation in several insect orders has facilitated their survival in various hostile environments. The subsequent use of monoamines and their N-acetylated metabolites in forming the sclerotized cuticle, which reduces water loss could be a partial explanation [16]. The presence of NAT and NAS in the neural tissues of the fly Drosophila has been known for over two decades.
Furthermore, the presence of the enzyme HIOMT, which is involved in melatonin production from NAS, has recently been suspected in insects. Antibodies produced against the pineal enzyme HIOMT, have been found to bind cockroach cells, indicating an enzymatic feature of the vertebrate pineal gland [17].
EFFECTS OF SEROTONIN AND MELATONIN
Effects of serotonin
In both protostomes and deuterostomes, the biogenic amine serotonin (5-HT) plays a critical role in regulating and influencing many physiological and behavioural processes. Its binding mediates serotoninŌĆÖs unique actions to membrane receptors and subsequent activation. The great majority of these receptors are members of the G-protein-coupled receptor superfamily. The discovery of 5-HT as a key neuromodulator required a collaborative effort from multiple research teams. In 1940, during the research of constricting factors in the blood that cause hypertension, a serum substance affecting vascular tone was separated and termed ŌĆ£serotoninŌĆØ [18,19].
5-HT was demonstrated to modulate appetite in several insect species. Neuromodulatory actions of 5-HT were shown to depress feeding in D. melanogaster, while decreased neuronal 5-HT levels increased appetite [20]. 5-HT injection in the hemolymph decreased feeding in another dipteran species, the flesh fly Neobellieria bullata [21]. 5-HT inhibits feeding when injected in the brain of honey bee, and when injected in the gut excited muscle contractions, although general elevation of 5-HT in the beeŌĆÖs hemolymph did not affect food intake [22]. Moreover, 5-HT increased fluid secretion from salivary glands in Aedes aegypti, the fly Calliphora vicina and the cockroach Periplaneta americana [23-25]. 5-HT acts as a diuretic hormone in Rhodnius prolixus, it is produced soon after the initiation of feeding and stimulates rapid tubule secretion [26,27].
5-HT also has thorough effects on some aspects of learning and memory and is thus a major player in modulating several insect behaviors. By inhibiting the serotonergic system in neurons of D. melanogaster, appetitive olfactory memory performance was considerably reduced [28]. Flies with genetically or pharmacologically reduced 5-HT levels in the brain also had a strongly reduced memory formation in a behavioral test wherein flies were trained to avoid a chamber position associated with high temperature [29]. In honey bees, both memory storage and retrieval were reduced when 5-HT was injected prior to conditioning [30-32]. In the desert locust Schistocerca gregaria, 5-HT and its downstream effector molecules were shown to induce gregarious, swarming behavior [33,34]. Fruit flies with genetically or pharmacologically elevated 5-HT levels showed higher fighting frequencies and more intense fighting than untreated flies [35]. In several dipteran species, manipulating serotonin levels with exogenous serotonin or serotonin agonists has been shown to affect feeding probability, modulate tarsal acceptance thresholds, and change meal size [36-39]. Exogenous serotonin injection decreases carbohydrate feeding in the grey flesh fly, N. bullata, and the queen blows fly, Phormia regina [36,39]. Although many aspects of fliesŌĆÖ feeding physiology have been studied, and many of the nervous pathways involved in feeding regulation have been identified, it is still unclear how and where serotonin influences feeding [40].
Effects of melatonin
Based on a review of the literature, it appears that the presence and synthesis of melatonin in photic and cerebral regions are a common trait of all invertebrates. Melatonin is present in the brain of Musca autumnalis changes regularly, with peak values seen during the dark period of the day-night cycle [41]. NAT activity of roughly 20 nmol/brain/hour in the cockroach brain and/or optic lobe, which is equivalent to that seen in chicken pineal glands at night. NAT does not appear to fluctuate regularly in the cockroach, whereas melatonin concentrations appear to be high at night and low throughout the day [42].
It has now been established that the pineal gland is involved in the transmission of photoperiodic information in vertebrates via the daily pattern of melatonin release. Invertebrates and vertebrates use photoperiodic variations as a time trigger to initiate physiological processes such as reproduction and diapause. Melatonin and its precursors or synthesizing enzymes are now well established in many organs implicated in photoreceptive processes or circadian pacemaking in both vertebrates and invertebrates.
In insects, melatonin synthesis and release follow a circadian rhythm influenced by light-dark cycles in the environment, just like in vertebrates. Insects use photoperiods as a time cue to start postembryonic processes such moulting, eclosion, and diapauses. This means they can tell the difference between long and short days.
Melatonin levels in insect species like M. autumnalis [41], D. melanogaster [43], L. migratoria [3], Gryllus bimaculatus [44], and Ischnura verticalis and Enallagma civile [45], R. prolixus [46], Ischnura graellsii, and Oedipoda caerulescens [47] or Apis mellifera [48] show a circadian rhythm with a scotophase peak typical of vertebrate melatonin production. The brain and optic lobes of the cockroach P. americana were described the same [42].
Even though melatonin is found in insects [3,49] and is thought to have physiological functions [50-53], the short-day mimicking effect was only detected in aphids [51]. Melatonin therapy delayed the commencement of oviposition in Pyrrhocoris apterus [50].
Insects benefit from exogenous melatonin because it activates the antioxidant defense system in response to a toxic insult. The effects of melatonin and its precursor serotonin (5-hydroxytryptamine) on the release of prothoracicotropic hormone (PTTH) from the brain of the studied cockroach species in vitro provided evidence for the first time that melatonin functions as a releaser of this glandotropic neuropeptide in this insect [54].