INTRODUCTION
Globally, preterm birth is the leading cause of neonatal death and child deaths <5 years [1]. In 2015, preterm birth alone responsible for 35.5% of newborn deaths and 17.8% of death age under 5 years [2]. The burden of preterm birth is highly significant in developing countries (18%) than the developed nations (5%) [1]. Exceptionally, the United States is one of the top 10 countries with the highest proportion of preterm births [1]. Although recent development in neonatal care has significantly improved, prematurely born babies still have a higher risk for long-term morbidities, such as neurodevelopmental disorder, cognitive impairments, etc. [3,4]. Therefore, the development of preventive measures for preterm birth; and novel treatment strategy for premature infants are urgently needed.
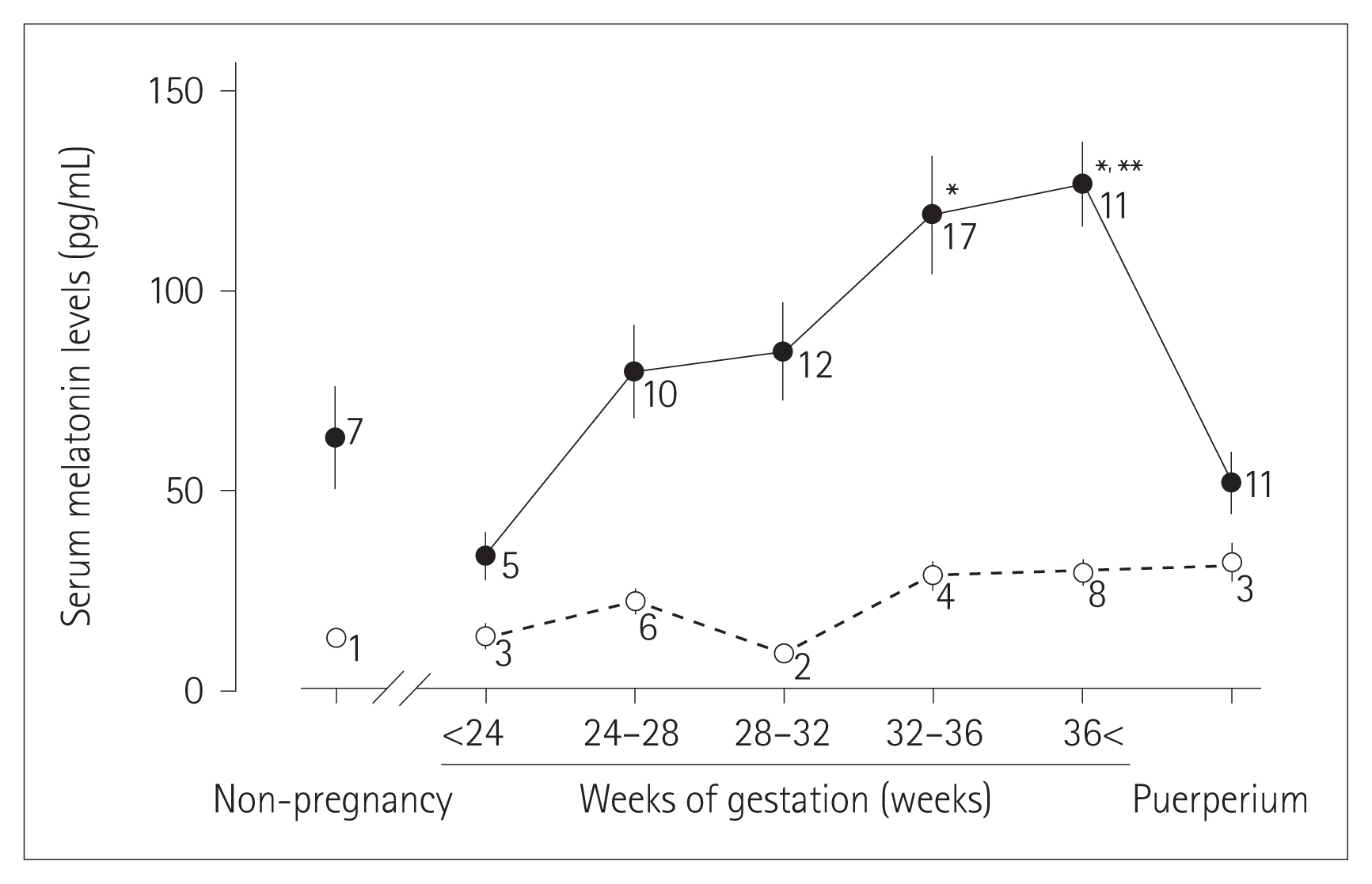
Melatonin (5-methoxy-N-acetyltryptamine) is a multifunctional hormone known for its role in numerous psychological and physiological responses including biological rhythms, endocrine regulation, antioxidant activity, sleep maintenance, and immune responses [5ŌĆō10]. A successful pregnancy requires coordination from all the physiological parameters between mother and fetus. Alteration of these crucial features may result in preterm births [11ŌĆō16]. Evidence suggests that maternal melatonin level is essential for fetal development and differential level of serum melatonin in pregnant and non-pregnant women has been reported (Figure 1) [17,18]. Okatani et al. [19] reported that melatonin able to cross the placental barrier, indicating maternal melatonin responsible for influencing the internal rhythms of the offspring. Disruption of internal melatonin rhythms may lead to preterm birth [20]. Most recently, one study concluded that night-shift working and continuous exposure to artificial light at unconventional times might compromise the success of full-term birth and the health of newborn infants [21]. Conversely, melatonin supplementation during pregnancy significantly increased gestation length and birth weight [22]. Furthermore, during pregnancy, melatonin treatment decreases preterm birth and perinatal brain injury [23]. Besides, melatonin intervention can also reduce brain damage in preterm neonates [24].
Melatonin had already shown its therapeutic potential for sleep disorders, jet lag, or shift work [25,26]. Generally, the action of melatonin is determined by the nocturnal secretion pattern. For instance, melatonin is involved in the regulation of sleep in diurnal species, which is associated with the length of night, or several metabolic processes associated with energy balance and seasonal control of photoperiodic functions. The external changes in the photoperiod can directly influence the metabolism in various species of vertebrates [6,27]. The artificial light at night and skyglow have been demonstrated to disturb the biological rhythm by influencing the secretion of melatonin which may disturb the pregnancy [18,27,28]. In this review, firstly, we explored the pre-clinical and clinical evidence to extend our knowledge on the role of melatonin during pregnancy. It is hypothesized that melatonin supplementation may improve the developmental and cognitive disorders in preterm infants. Therefore, secondly, we discuss the recent progress regarding the favorable effects of exogenous melatonin on preterm birth, with particular emphasis on respiratory diseases and brain injury. Finally, we discussed how chronodisruption is associated with a high risk of preterm birth.
MELATONIN BIOSYNTHESIS
Melatonin can be considered as ancient as it seems to have appeared around 2.5ŌĆō3.0 billion years ago in photosynthetic cyanobacteria [29]. Moreover, melatonin has a highly conserved pattern of biosynthesis. It is high during night and nadir at day time; therefore, it is considered the most important ŌĆ£chronobioticŌĆØ molecule and endogenous neurohormonal signal of darkness [5,10,30]. The chronobiotic molecule melatonin, an indoleamine, is mostly synthesized in the pineal gland of vertebrates. Information from studies of the last few years has enriched us about the presence and synthesis of melatonin in the retina, gut, ovary, and brain [31ŌĆō35]. The synthesis of melatonin is very much conserved and classically involves four enzymatic processes using the amino acid tryptophan as the initial substrate. 5-hydroxytryptophan is produced from tryptophan by tryptophan hydroxylase. The serotonin or 5-hydroxytryptamine (5-HT) is the next product by the action of 5-hydroxytryptophan decarboxylase. The 5-HT is acetylated to N-acetylserotonin by arylalkylamine N-acetyltransferase. Finally, methylation of the last product by hydroxyindole-O-methyltransferase forms the melatonin [36ŌĆō39].
Local melatonin synthesis has been reported in other tissues and cells in mammals, which has a minor effect on plasma or cerebrospinal fluid (CSF) levels of melatonin. For instance, melatonin is synthesized in the retina in a circadian manner and participates in the regulation of retinal physiology, including retinal light sensitivity during the night [40]. The gastrointestinal epithelium also produces melatonin from its precursor serotonin, which is thought to come from enterochromaffin cells. In the gut, melatonin seems to participate in the regulation of intestinal motility, immune system responses, ion transportation in the lower gut, and the release of peptides involved in energy balance such as peptide YY [41].
FUNCTION OF MELATONIN IN PREGNANCY
Melatonin decreases in conditions associated with serious outcomes for the fetus and seems to be involved in preeclampsia and intrauterine growth restriction [11]. Melatonin treatment during human normal or abnormal pregnancy has been studied for a large range of conditions and at different times during the gestational period. Considering the ethical issues, it is more difficult to study a normally occurring pregnancy, than an in vitro fertilization (IVF) one. Melatonin administration started before IVF-cycles, continued during pregnancy, and was associated with improved pregnancy outcomes [8]. Melatonin receptors are widespread in the human fetus from early fetal development. Also, it appears that the fetusesŌĆÖ sleep patterns develop in late pregnancy, melatonin being the regulating factor. A normal sleep pattern is involved in neurodevelopment and there is solid evidence that melatonin is involved in fetal neuroprotection. Thus, the influence of melatonin on the developing human fetus may not be limited to entertaining the circadian rhythmicity. Melatonin research showed that it plays an important role in pregnancy and parturition. The passage of maternal melatonin through the placenta exposes the fetus to a daily melatonin rhythm of low concentrations during the day and high concentrations at night. Therefore, first, melatonin is involved in inducing a circadian manner of functioning in fetal organs.
The ability of melatonin to promote embryo development in different species has been reported. When mouse embryos were cultured in a medium containing melatonin, increased blastocyst development rates were observed [17]. Melatonin has a beneficial role in the in vitro development of rodent embryos found in the 2-cell stage [19] and helped the maturation of bovine blastocysts [20]. Suppression of maternal plasma melatonin circadian rhythm by continuous light exposure during the second half of gestation showed several effects on fetal development. First, it induced intrauterine growth retardation. Second, in the fetal adrenal in vivo it markedly affected the mRNA expression level of the clock genes and clock-controlled genes, as well as lowered the content and modified the rhythm of corticosterone. Third, an altered in vitro fetal adrenal response to adrenocorticotropic hormone (ACTH) of both, corticosterone production and relative expression of clock genes and steroidogenic genes was observed. All these changes were reversed when the mother received a daily dose of melatonin during the subjective night [21].
Torres-Farfan et al. [22] reported that maternal melatonin decreased cortisol production in the fetal adrenal gland of the capuchin monkey. In another study on sheep, they found that melatonin had direct inhibitory effects on the noradrenalin-stimulated fetal cerebral artery contraction, the release of glycerol by brown adipose tissue, and on ACTH-induced secretion of cortisol by the fetal adrenal gland. Low levels or a lack of a circadian rhythm of the fetal corticosterone may be the cause of the intrauterine growth retardation that has been previously reported. The lack of maternal melatonin (induced by pinealectomy) during the early stages of gestation was found to disrupt the drinking behavior of rat pups, an effect reversed by the administration of exogenous melatonin to the dam [23]. Melatonin is important in normal placental development and function, a function supported by the placenta melatonin receptors expression during early pregnancy [24]. Whereas melatonin synthesis is classically considered to take place in the cytoplasm, in the past 5 years, non-rhythmic melatonin synthesis has been reported to occur in isolated mitochondria from mouse brain cells and oocytes, as well as in plant mitochondria and chloroplasts. The full meaning of this intriguing finding remains to be clarified, including the question of the effect of mitochondrial melatonin synthesis in comparison to pineal melatonin synthesis.
ROLE OF MELATONIN IN PRETERM BIRTH
It is hypothesized that deprivation of long-term endogenous melatonin levels may link with neurological and developmental disabilities in premature neonates. In fact, after full-term birth, the neonate unable to generate melatonin for the first 8ŌĆō16 weeksŌĆöa phase is called transient melatonin deficiency [1ŌĆō5]. Although previous studies showed that the suprachiasmatic nucleus and the pineal gland had appeared to mature in early fetal life [3], however, the neurological circuitry that controls these structures remain immature [6,7]. Therefore, in both preterm and full-term neonates (absence of maternal melatonin), the uprising of the circadian pattern is crucially dependent on the neurodevelopmental status rather than the home environment [8]. Prematurity itself is negatively associated with the maturation of the neurological network, which controls melatonin secretion [9]. Evidence suggests that the impairment for the development of the neurological network delayed the onset of pineal melatonin secretion [2,5]. Consequently, in the preterm neonates, the deficiency of endogenous melatonin levels is significantly longer than the full-term neonate [1,2,4,5]. Extensive research has shown that abnormal brain, lungs, and respiratory system development most frequently occur in the premature infant. Data from the previous study suggest that this may be associated with a prolonged deficiency of pineal melatonin levels, which may be restored with melatonin supplementation [10]. In this section, we discuss the recent progress regarding the beneficial effects of melatonin on preterm birth, with particular emphasis on respiratory diseases and brain injury.
The pivotal role of melatonin in respiratory diseases
Bronchopulmonary dysplasia (BPD), a chronic lung disease that requires respiratory support at birth, is most frequently appeared in preterm infants whose gestation age <29 weeks [11]. Pathogenesis of BPD has been attributed to multiple factors, including prematurity of lung tissue, superabundant of the inflammatory state, and disruption of the repairing process following injury [12ŌĆō14]. In light of the scientific advances in the field of neonatology, newer and effective perinatal strategies have come out to fight against BPD, for instance, new formulations of surfactant (makes breathing easier), application of corticosteroids, and non-invasive ventilation [15]. Despite this, BPD remains one of the most common complications associated with prematurity [16]. Most recently, one randomized clinical trial had conducted on 100 premature neonates who had been suffering from severe respiratory distress syndrome (RDS) by mechanical ventilation [17]. In this study, the premature neonates were allocated into two groups: group one population received melatonin supplementation and group two without melatonin supplementation. The results of this study revealed that the melatonin treated group significantly declined in the length of incubator stay of neonates and markedly decreased in the development of established cases of BPD, in comparison to that of premature neonates who did not take melatonin [17], indicating melatonin may use as an adjuvant therapy for the prevention of BPD in premature neonates. In another clinical trial, it has been reported that the inflammatory markers, including interleukin-6 (IL-6), interleukin-8 (IL-8), and tumor necrosis factor-╬▒ (TNF-╬▒), were significantly higher at one day, three days, and at seven days in preterm neonates with RDS, whereas melatonin treatment can block these inflammatory components and likely reduce the severity of RDS in preterm newborns [18]. Furthermore, melatonin treatment in preterm infants with RDS not only decreased inflammatory markers but also reduced nitrite/nitrate levels in comparison with the placebo-treated group [19]. In BPD, immature lung recruits inflammatory cells and overexpress inflammatory molecules, which are considered the mainstem pathologic pathway to tissue damage [20]. Based on pathological consideration, melatonin may exert beneficial effects in the incidence of BPD, particularly in preterm newborns. However, the data from the aforementioned studies cannot support the beneficial effects of melatonin for long-term respiratory outcomes in preterm infants, which need further investigation. In hyperoxia-induced BPD animals, the lung myeloperoxidase and nitrite/nitrate ratio were decreased through melatonin intervention [21]. Moreover, melatonin treated group significantly increased antioxidant enzymatic activities, including glutathione peroxidase, superoxide dismutase, and catalase activity [42]. These findings indicate melatonin contributes to lung architecture development and protects lung during the critical processes of alveolarization following preterm birth.
Neuroprotective role of melatonin in brain injury
Recent developments in neonatology have focused on fetal brain injury, which is considered one of the leading causes of morbidity and mortality in preterm infants [22]. Intraventricular hemorrhage (IVH, where CSF is produced) and white matter abnormalities frequently appear in preterm infants, resulting in neurosensory and cognitive disabilities [23ŌĆō25]. The cellular and molecular mechanisms of preterm infantsŌĆÖ brain injury to be complex and multifactorial. Evidence suggests that oxidative stress is among the most important risk factors for brain damage in preterm newborns [24]. Previous findings suggest that neuronal cells are more vulnerable to hypoxic-ischemic injury in the preterm brain than in term neonates [24], particularly related to high levels of reactive oxygen species that are responsible for significant neuronal damage [43]. Relatively the immature brains are more prone to oxidative stress than the matured brain because the immature brain contains a higher proportion of blood vessels, and their cortex poorly developed when compared with mature ones [44]. Particularly in preterm neonates, morbidity and mortality are significantly influenced by their ability to maintain physiological homeostasis [45]. Interestingly, disruption of the antioxidant defense system most often appeared in preterm infants, consequently, they are unable to counteract the detrimental effects of free radicals [45]. Data from several pre-clinical studies suggest that melatonin exerts neuroprotective effects in the preterm infant brain. For example, in the umbilical cord occlusion induced asphyxia, the oxidative stress was decreased with melatonin treatment, in both preterm and near-term fetal sheep [22]. Chowdhury and Maitra [32] also reported that melatonin reduced cell death in the fetal brain following intrauterine asphyxia by decreasing inflammation in preterm fetal sheep.
One recent study demonstrated that the administration of melatonin before and after hypoxia in immature rats has markedly attenuated brain injury and improved behavioral asymmetry, suggesting melatonin would be a potentially safe drug to perinatal brain damage in humans [46]. To better understand the difference between pharmacokinetic parameters of melatonin in preterm infants and adults, Merchant et al. [34] had compared the preterm infant pharmacokinetic profile (PP) with extrapolating adult pharmacokinetic data. The principal findings of this study demonstrated that the PP of melatonin in preterm neonates differs from that of adults. Therefore, for preterm infants, the melatonin dosage cannot be extrapolated from adult studies. These results are similar to those reported by Carloni et al. [35]. Most recently, in preterm fetal sheep, Yawno et al. [24] examined whether melatonin could recover white matter brain development when administered after an acute hypoxic-ischemic insult. It is apparent from this study that hypoxic-ischemia causes a notable increase in apoptosis, microglial activation, and oxidative stress in white matter areas. Whereas melatonin treatment significantly reduced apoptosis, inflammation, and oxidative stress following hypoxic-ischemia [36].
PRETERM BIRTH IN SHIFT WORKERS AND INSOMNIA
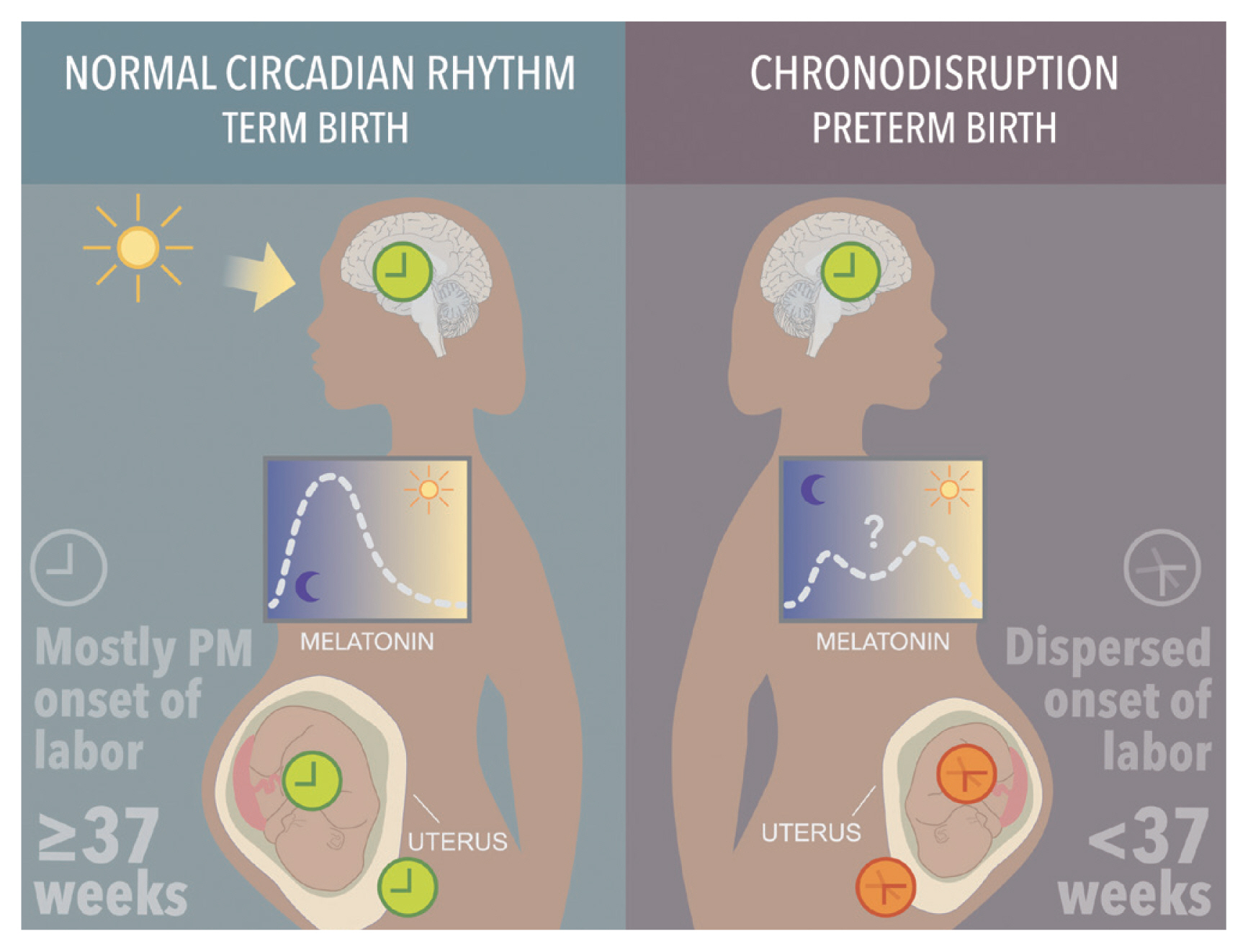
Chronodisruption leads to reproductive dysfunction and a low level of melatonin appears to be a key contributor to preterm birth and associated diseases that develop in adult life (the concept of fetal programming) (Figure 2) [47]. Women working an evening shift or a night shift, or who are trans meridian traveling, report an increased or decreased menstrual cycle length, changes in the duration and amount of menstrual bleeding, and dysmenorrhea [48ŌĆō50]. These symptoms are not subjective, as they correlate with changes in the hormonal profile of ovarian and pituitary origin, such as a high follicular stimulating hormone (FSH) and prolonged follicular stage of the ovarian cycle [50]. The working environment also affects pregnancy outcomes. The shift work while pregnant is also associated with a high risk of prematurity and/ or low for gestational age babies, spontaneous abortion, and subfecundity [51,52]. Shift work and jet lag effects on health may be explained by the secondary reduction of the total amount of sleep. FSH concentration in women who sleep less than 8 h/night lowers by 1/5 compared to women with longer sleep duration [53]. Total or partial sleep deprivation increases luteinizing hormone (LH) amplitude, estradiol, and FSH concentrations in normal cycling women, and increased estrogen is associated with a high risk of breast cancer. Cumulatively, shift work, or poor sleep may become a crucial factor in preterm birth.
THERAPEUTIC POTENTIAL OF MELATONIN IN PRETERM BIRTH
Preterm birth (birth before 37-week gestation) nowadays is a serious health issue worldwide. Over 13 million babies were born in the preterm condition in 2005 and more than 1 million infants dying every year due to preterm born. Those who survived face an increased risk of morbidities, such as cerebral palsy, autism, schizophrenia, blindness, and hearing loss [54]. In recent days many advances have been made in the medical field of obstetrics and neonatology, but the rate of premature delivery has not decreased yet over the past 20 to 30 years. To date, there is no efficient therapy to prevent preterm labor and increase the survival of offspring as no precise pathways have been discovered for this preterm parturition. Despite that, from some epidemiological and experimental data, it is revealed that maternal infection and inflammation are the key factors in preterm birth. Approximately 40% of all premature deliveries occur to mothers who have an intrauterine infection [55]. Working outside of regular work, like shift working, night working highly prevalent in both industrialized and post-industrialized societies. Particularly health care professionals, more are women and many of them are of reproductive age which can have a hazardous effect on pregnancy and fetal development [52]. The circadian rhythm aligns many physiological and behavioral rhythms in the mother according to that fetus also synchronizes its oscillating circadian rhythms and this tightly coordinated mechanism helps in the developmental processes [56,57]. During night work circadian rhythm of the motherŌĆÖs body is disrupted due to the exposure to light [58]. This type of repeated and consecutive changes in maternal circadian rhythm affects fetal development as fetuses failed to synchronize their oscillating circadian rhythms with mother. Exposure to light during the dark period suppresses the amplitude of melatonin secretion, which might further affect the circadian rhythms of the mother body. Melatonin possess antioxidant, a free radical scavenger and anti-inflammatory properties which indicate it have a role in reducing oxidative stress [59ŌĆō61]. It is evident that the melatonin is elevated during pregnancy and it can cross the placenta, blood-brain barrier of fetus without showing any risk for mother and fetus [62,63]. Bacterial infection in the uterus, placenta, and fetal tissues stimulates cytokine and chemokine responses by leukocytes and other cells which leads to the release of prostaglandins (PGs), nitric oxide (NO), and matrix metalloproteinases [55]. It has been reported that PGs, NO, and TNF-╬▒ are mainly responsible for preterm labor in several rodent models as well as in humans [64ŌĆō66]. A high level of plasma melatonin during late pregnancy helps in gestation maintenance by stimulating progesterone production and inhibiting PG synthesis and uterine contractility [67]. In mice, melatonin protects from lipopolysaccharide (LPS)-induced fetal death, intrauterine growth restriction, and cellular stress in the placenta [68] while other research demonstrated that maternally administered melatonin differentially regulates LPS-induced pro-inflammatory and anti-inflammatory cytokine levels in maternal serum, amniotic fluid, fetal liver, and fetal brain [69].
It has been reported that increased NO, PG, and cytokine plays a significant role in preterm delivery. Whatever, meloxicam (a COX-2 inhibitor), aminoguanidine (an iNOS inhibitor), etanercept (a competitive inhibitor of TNF-╬▒) prevent inflammation-induced preterm delivery [65,70], besides that the impairment of PG/COX-2 system, decrease in NO levels, diminishing cytokine production or combination of these treatments may be a therapeutic strategy to prevent preterm labor [67]. In that way melatonin could be a promising resource in the managing of preterm labor as it decreases PGs, NO, IL-1╬▓, IL-6, COX-2, and TNF-╬▒ levels and can regulate the circadian rhythm in mice uterus and other mammals [71,72]. Besides that, melatonin is a very safe compound for human use, may be considered as a novel tocolytic agent to treatment against inflammation-associated preterm birth in clinical trials [23].
CONCLUSIONS
In conclusion, it may be claimed that melatonin may play a pivotal role in reducing preterm birth. While the clinical applicability of melatonin is yet to be elucidated, it remains a very promising candidate in avoiding preterm birth or the diseases associated with preterm birth. More clinical studies, pharmacokinetics, and pharmacodynamic analysis of melatonin in pregnant women, placenta, and the embryo are required to further confirm the role and potency of melatonin in reducing preterm birth.