


Withanolide-A Accelerates Re-entrainment of Circadian Locomotor Activity Rhythm and Preserves Spatial Memory in Mice Following Acute Phase Inversion of Light-Dark Cycles
-
Trisha Tiwari1
 , Priyoneel Basu1,2, Muniyandi Singaravel1
, Priyoneel Basu1,2, Muniyandi Singaravel1
- Received June 13, 2022 Revised August 28, 2022 Accepted August 30, 2022
- ABSTRACT
-
- Objective
- Circadian rhythms are present almost in all living organisms and synchronized with the zeitgebers (time givers), especially light-dark (LD) cycles. The jetlag and rotating shift work schedules lead to desynchronization of the circadian clock with zeitgebers which may cause various health problems. Studies have shown that withanolide-A (WA) is a potent nootropic and neuroprotective agent found in Withania somnifera. Thus, the objective was to study the effect of WA on the circadian locomotor activity rhythm following the phase inversion of 12:12 h LD cycle in mice and its impact on the spatial memory in mice.
- Methods
- Adult male mice were housed in the chronocubicle and recorded circadian locomotor activity rhythm by using ClockLab setup. The experimental mice were administered with WA (10 μmol/kg/day) for 13 consecutive days at about ZT15 and then evoked an acute phase inversion of 12:12 h light-dark cycle. Further, the Barnes maze test was performed to check the effect of WA on spatial memory following the phase inversion.
- Results
- The results showed that WA-treated group took significantly fewer transient cycles (4.66+0.66) for re-entrainment than the control (9.5+0.43) and vehicle-treated (8.33+1.28) mice following the phase inversion. Similarly, WA-treated mice traveled less distance to reach the target zone in the Barnes maze compared to vehicle and control groups.
- Conclusion
- These results suggest that the WA may act as chronobiotic and neuroprotective agent since it decreased the transient cycles for re-entrainment and retaining spatial memory following the phase inversion.
- INTRODUCTION
- INTRODUCTION
Circadian rhythms exist ubiquitously in nature and enable adaptation to appropriate temporal niches through the regulation of physiological variables, including heart rate, hormones, neurotransmitters, biochemical pathways, and learning and memory, among many other processes, via interaction with rhythmically transcribing genes (clock genes) [1]. The photic or non-photic cues that modulate the circadian rhythm of organisms are called zeitgebers (time givers). The organisms consist of the circadian clock that can both respond to rhythmic external stimuli and maintain an endogenous rhythm and anticipate environmental changes [2]. In contemporary lifestyles, frequent trans-meridian travel, rotating shift work schedules, and irregular sleep patterns result in a lack of synchronization between the endogenous circadian timing system and the external environment [3]. This leads to multiple health issues including cognitive deficits [4], metabolic disturbances [5-7], cancer [8], increased mortality [9,10], and many others. Abrupt changes in the light-dark cycle cause desynchrony between the entrained state of the endogenous clock and external input (stimuli). Such desynchrony occurs during shift work, when the individual works in a different light-dark schedule than the natural one, or during trans-meridian flight where several time zones are rapidly crossed, causing jetlag [10]. A person traveling from the USA to India thus has to readjust to a light-dark schedule that is approximately 12 h ahead of the previous schedule. This physiological readjustment takes a few days to complete. During this period, onsets and offsets of activity shift gradually in a series of steps called transient cycles. Appropriately timed light exposure (photic means) or the administration of a chemical compound (non-photic means) [11] can hasten re-entrainment.India has many plants containing clinically active components. Withania somnifera or ashwagandha, also known as Indian ginseng, has been used in Indian traditional medicine for several centuries and is an established tonic and nootropic agent [12]. Ashwagandha is a source of many active compounds including withaferins, withanone, withanolides, etc. Withanolide-A (WA) has a neuroprotective effect in induced neuroinflammation [13]. It has recently been proved that withaferin-A can lengthen the circadian period, upregulate Bmal1, and act as an inverse agonist of RORα [14], thus exhibiting a potential to act as a chronotherapeutic agent. Previously, Withania extracts have been shown to restore Sirt1 and Nrf2 rhythms [15] during aging.Jetlag is known to cause cognitive deficits during the period of re-entrainment [16], affecting neural precursor cell proliferation and inhibiting functional brain plasticity. Agents that help ameliorate the ill effects of jetlag, such as melatonin, may also help to prevent cognitive deficits [17].Based on the information outlined above, we hypothesized that WA, a steroidal lactone found in Withania, may act as a chronotherapeutic as well as a neuroprotective agent during abrupt changes in light-dark cycles which causes circadian misalignments, such as jetlag or inversion of the circadian phase. Withanolides are C28 steroids that occur naturally. Their structure is based on an ergostane skeleton in which C-26 and C-22, or C-26 and C-23, are oxidized to form either delta lactone (Type A) or gamma lactone (Type B), respectively [18]. The axonal outgrowth of normal cortical neurons after WA treatment has been reported [12]. However, to the best of our knowledge, the putative dual chronotherapeutic and neuroprotective effect of WA has not been explored.Therefore, the objective was to study the effect of WA on the circadian rhythm of mice subjected to an acute phase inversion of the 12:12 h light-dark schedule. Further the focus was to elucidate the neuroprotective effect of WA on spatial memory of mice exposed to phase inversion with help of the Barnes maze behavioural test.
- METHODS
- METHODS
- Subjects
- Subjects
Adult male mice, Mus musculus (AKR strain) of about three months old were used for this experiment. The mice were maintained in the animal house under the 12:12 h light-dark cycle with the temperature of 25°C±2°C. The food pellets and water were provided ad libitum. An electronic timer (Larsen and Toubro, Mumbai, India) was used to maintain the light-dark cycle. The light intensity at the cage level was about 150 lx during the light phase and 0 lx during the dark phase. A red-filtered torch light (<1 lx) was used during dark hours for maintenance, for which visits were made at random times. The Institutional Animal Ethical Committee norms of the University (Dean/13–14/CAEC/185), the recommended ethical considerations for biological rhythm research [19], and the specified maintenance and analysis protocols by Jud et al. [20] were followed for this investigation.- Experimental groups
- Experimental groups
The animals were divided into two sets. Set-1 included 18 adult mice, divided into three groups: 1) a control group (n=6) exposed to acute-phase inversion of light-dark schedule; 2) a vehicle-treated group (n=6) (0.5% gum arabic, HiMedia, Mumbai, India), which was exposed to an acute-phase inversion of light-dark schedule and treated with a single dose of the vehicle per day; and 3) a WA-treated group (n=6, 10 μmol/kg/day, Cayman, Ann Arbor, MI, USA) given a single oral dose each day and exposed to an acute phase inversion of light-dark schedule at the end of the dosing period. All the doses were administered orally for 13 consecutive days at approximately zeitgeber time (ZT) 15 and the light-dark cycle was inverted on the day following the last dose.The Set-2 consisted of 15 mice divided into three groups for Barnes maze behavioural test: 1) the control group (n=5) was exposed to phase inversion only; 2) the vehicle group (n=5) was treated with a single dose of the vehicle per day; and 3) the experimental group (n=5) administered with a single oral dose of WA (n=5) (10 μmol/kg/day) each day, for 13 days. Each of these groups was exposed to an acute-phase inversion of the light-dark schedule. A separate set of mice (n=6) was retained for performing a Barnes maze test without undergoing jetlag or receiving WA but maintained in conditions similar to the other groups (untreated group).- Measures
- Measures
Simulation of acute phase inversion and locomotor activity rhythm recording
Simulation of acute phase inversion and locomotor activity rhythm recording
The chronocubicle was maintained with a 12:12 h light-dark schedule (06:00 h to 18:00 h light, 18:00 h to 06:00 h dark), and after 10 days of acclimation to the light-dark schedule and baseline wheel recording, oral administration of doses was given for 13 consecutive days to the groups described above; vehicle group (0.5% gum arabic solution) and WA group (10 μmol/kg/day) at around ZT15. The following day, the light-dark schedule was inverted (18:00 h to 06:00 h light, 06:00 h to 18:00 h dark), and this was maintained till the mice were re-entrained to the new light-dark schedule. The ClockLab (version 2.63; Actimetrics, https://actimetrics.com/products/clocklab/) setup was used to record and analyze the voluntary wheel-running activity. Mice were allowed unlimited voluntary access to running wheels (11.5 cm diameter) inside individual living chambers (36 cm × 20 cm × 14 cm). The 12:12 h light-dark schedule was maintained and inverted using an electronic timer. The untreated group was maintained in an adjoining chamber under the 12:12 h light-dark schedule and similar handling and maintenance procedures. The wheel revolutions as recorded by magnetic reed-relay switches were transmitted to a dedicated computer via an ACT-553 7-channel breakout box and a CL-200 56-channel interface. Data were recorded in 6-minute (0.1 h) interval bins as counts/min and analyzed using the ClockLab (version 2.63) software on the MATLAB platform (MathWorks, Natick, MA, USA).Spatial memory test
Spatial memory test
In brief, the Barnes maze was used for the spatial memory test, comprised of three phases: habituation, training, and probe trial [21]. In the habituation phase (1st day), mice were allowed to explore the maze apparatus freely for 180 s. The training was given for 3 consecutive days; each mouse was given 3 trials of 180 s each day, having an inter-trial difference of 15 min. A probe test (90 s) was done after 48 h of inversion of the light-dark schedule to test the retention power of mice that were trained during the training phase before the phase inversion. The training and testing procedures were performed during the active period of mice, around ZT15. After each trial, the maze was wiped with 70% ethanol and allowed to dry to remove any olfactory cues. Barnes maze behavior test was performed for all three groups; control, vehicle, and treated, and separately for the untreated group and the data were analyzed by using SMART Video Tracking System (Panlab, Harvard Apparatus, Holliston, MA, USA).
- Statistical analyses
- Statistical analyses
Circadian behavioral rhythms were expressed as double plotted actograms in 6 min bins, and the phases of activity onset were estimated using ClockLab software (version 2.63). The transient cycles (number of days taken to get re-entrained to the new light-dark schedule) were compared between the control, vehicle, and WA-treated groups. The number of transient days was counted manually (marked by arrows on representative actograms of each group) for each mouse of their respective group. Further, their mean was taken and tested for any statistical difference between the three experimental groups, by Student’s t-test and one-way analysis of variance (ANOVA) by Microsoft Excel 2021. Actograms were analyzed using ClockLab software and the data were analyzed using Microsoft Excel.
- RESULTS
- RESULTS
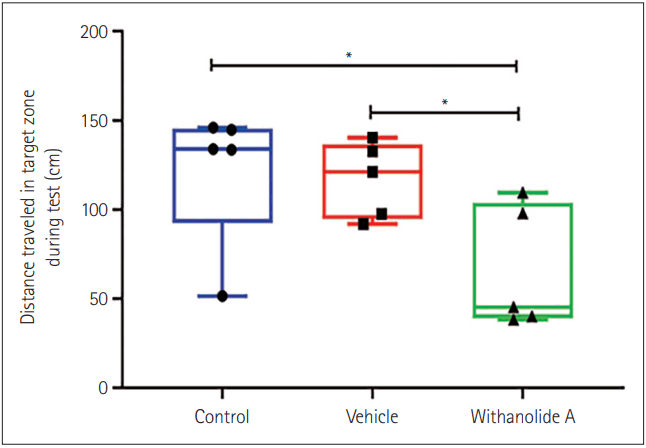
WA administered at around ZT15 caused faster entrainment in mice compared to control and vehicle groups, as evident in the actograms (Figure 1). The number of transient days required to entrain to acute-phase inversion of 12:12 h light-dark schedule in the WA treated group was significantly lesser (p<0.05, one-way ANOVA). Mice from the control group took more days (9.5±0.43 days) compared to the vehicle (8.33±1.28 days) and WA-treated group (4.66±0.66 days) (Figure 2). The WA-treated group thus re-entrained faster than the control and vehicle-treated groups (F[2,15]=16.42, p<0.05). The average day-wise onset of activity before and after phase inversion of the light-dark schedule was also analyzed in all three groups (Figure 3) and shows clearly the number of transient days is lesser in the WA treated group.The Barnes maze test was performed to investigate the effect of WA on spatial memory retention of mice that were given pre-doses and training before the light-dark schedule inversion. The successive day-wise improvement in learning in all groups was analyzed as the distance traveled in the target zone (in which the escape chamber was located) decreases on successive days (Figure 4). Mice not exposed to acute phase inversion traveled less distance in the target zone and found the escape hole faster than exposed mice (67.8±8.42 cm vs. 121.89±17.80 cm) and vehicle-treated mice (67.8±8.42 cm vs. 116.7±9.52 cm) (p<0.05, Student’s t-test). Post-exposure, WA-treated mice traveled less distance in the Barnes maze test (66.17±15.45 cm) compared to vehicle and control groups after the phase inversion (Figures 4 and 5). A significant difference was seen in the distance traveled in the target zone during the probe test performed after phase inversion of the light-dark schedule (F[2,12]=4.39, p<0.05) between WA-treated, vehicle, and control groups. This shows that WA-treated group has better retention power than the other two groups.
- DISCUSSION
- DISCUSSION
Our study demonstrated the dual potential of WA in both hastening re-entrainment to acute phase inversion of the zeitgeber/light-dark cycle and a possible neuroprotective effect during shift work/jetlag. It is well-known that jetlag produces cognitive deficits [4,22]. The phase inversion represents an extreme form of acute shift work/jetlag. Accordingly, in both control and vehicle-treated groups, performance in the Barnes maze after inducing acute phase inversion was impaired, compared to the group not exposed to phase inversion (untreated group) (graph not shown). WA pre-treated mice showed remarkable resistance to cognitive deficit compared to vehicle-treated and control groups when subjected to acute phase inversion (Figures 4 and 5). WA pre-treated animals achieved values close to those of untreated mice that had not faced phase inversion. A recent study showed that the administration of melatonin restores hippocampal neural precursor cell proliferation and prevents cognitive deficits in adult mice that were exposed to photoperiod alterations by shortening the dark period every third day by 6 hours for 3 weeks [17]. The treatment of WA shows similar results on spatial memory suggesting that WA may also be acting as a neuroprotective agent (Figure 5). There was marked inter-individual variation in the WA treated group, suggesting that a larger sample size may be necessary for further studies.WA also hastened re-entrainment to acute phase inversion (Figures 1-3). While the authors do not offer a mechanistic explanation at this point, this effect is probably through the interaction of WA with Bmal1, RORα, and other components of the molecular circadian clock [14]. It is possible that the neuroprotective effect of WA is partly due to faster re-entrainment, and hence less damage, and that these two phenomena are synergistically tied together.To the best of our knowledge, a study on the dual effects of WA on the acceleration of re-entrainment of the circadian clock and its neuroprotective effect on post-circadian rhythm inversion has not been carried out before. Similar to the WA effects on re-entrainment, both exogenous melatonin and scheduled feeding accelerated re-entrainment of the suprachiasmatic nuclei–driven general activity after a 6 h of phase advance of the light-dark cycle in rats [23]. Also, melatonin administration in males exposed to 8 h advanced jetlag, re-entrain faster [24]. The endogenous melatonin signal facilitates re-entrainment of the circadian system faster compared to mice which are deficient in melatonin or its receptors [25]. Even though WA is accelerating the re-entrainment of locomotor activity rhythm similar to melatonin, the mechanism of action is yet to be elucidated.In today’s paradigms, shift-work schedules and phase inversions are fast becoming the norm, rather than the exception, with approximately 20% of the world’s working population undertaking shift work [26]. Hence, chronotherapeutics that helps ameliorate the ill effects of shift work would be of use. WA, and by extension, Withania extract, with its dual benefits, and ease of administration, would be a helpful addition to the repertoire of agents that can help prevent the ill effects of shift work. There is scope for further investigation of this phenomenon at the molecular level.
- NOTES
- NOTES
-
Funding Statement This research work was supported by the Department of Science and Technology, India [SR/CSI/157/2012] to MS.
The authors have no potential conflicts of interest to disclose.
Availability of Data and Material The datasets generated or analyzed during the study are available from the corresponding author on reasonable request.
Author Contributions Conceptualization: Muniyandi Singaravel, Trisha Tiwari. Data curation: Trisha Tiwari. Formal analysis: Trisha Tiwari. Funding acquisition: Muniyandi Singaravel. Investigation: Trisha Tiwari, Muniyandi Singaravel. Methodology: Trisha Tiwari. Project administration: Muniyandi Singaravel. Resources: Muniyandi Singaravel. Supervision: Muniyandi Singaravel. Validation: Priyoneel Basu, Muniyandi Singaravel. Visualization: all authors. Writing—original draft: Trisha Tiwari, Priyoneel Basu. Writing—review & editing: Priyoneel Basu, Muniyandi Singaravel.
- ACKNOWLEDGMENTS
- ACKNOWLEDGMENTS
ICMR Senior Research Fellowship to TT [Award No.3/1/3/3/M/2019/NCD-1], Departmental facilities of DST-FIST, UGCCAS, ISLS-BHU, and IoE-BHU are acknowledged.
Figure 1.

Figure 2.
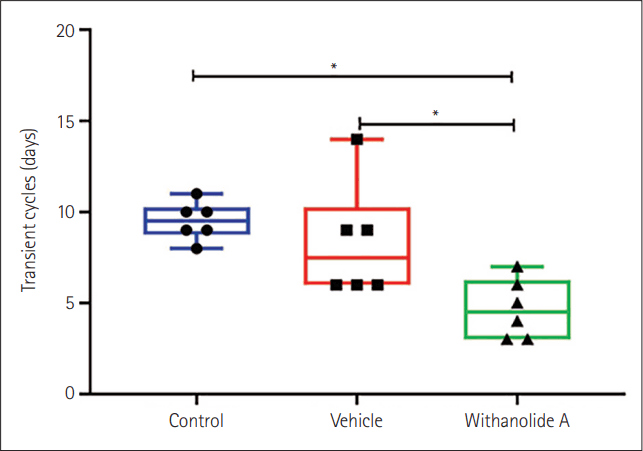
Figure 3.
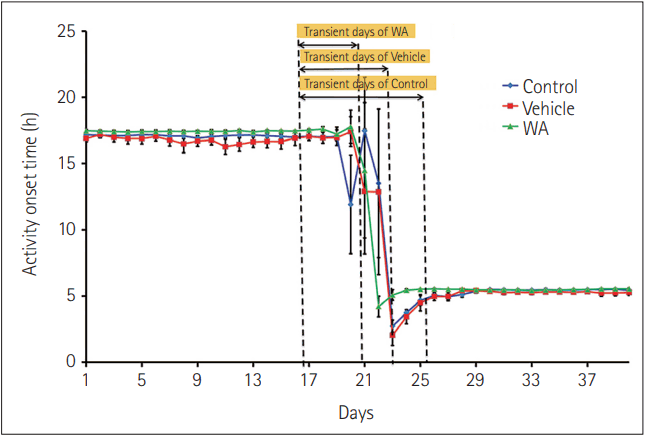
Figure 4.
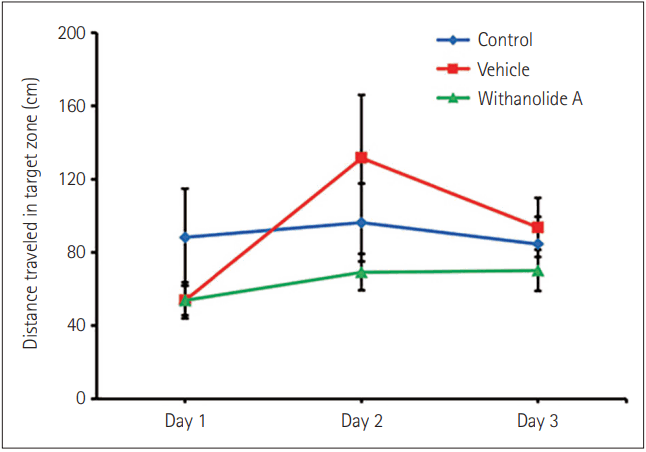
Figure 5.
- REFERENCES
- REFERENCES
- 1. DeCoursey PJ, Walker JK, Smith SA. A circadian pacemaker in free-living chipmunks: essential for survival? J Comp Physiol A 2000;186:169–180.
[Article] [PubMed]2. Sharma A, Tiwari S, Singaravel M. Circadian rhythm disruption: health consequences. Biol Rhythm Res 2016;47:191–213.
[Article]3. Kumar D, Basu P, Singaravel M. Variations in the rate and direction of reentrainment to acute simulated jet lag in the diurnal North Indian palm squirrel. Biol Rhythm Res 2014;45:447–454.
[Article]4. Gibson EM, Wang C, Tjho S, Khattar N, Kriegsfeld LJ. Experimental ‘jet lag’ inhibits adult neurogenesis and produces long-term cognitive deficits in female hamsters. PLoS One 2010;5:e15267.
[Article] [PubMed] [PMC]5. Karatsoreos IN, Bhagat S, Bloss EB, Morrison JH, McEwen BS. Disruption of circadian clocks has ramifications for metabolism, brain, and behavior. Proc Natl Acad Sci U S A 2011;108:1657–1662.
[Article] [PubMed] [PMC]6. Pati AK, Chandrawanshi A, Reinberg A. Shift work: consequences and management. Curr Sci 2001;81:32–52.7. Huang W, Ramsey KM, Marcheva B, Bass J. Circadian rhythms, sleep, and metabolism. J Clin Invest 2011;121:2133–2141.
[Article] [PubMed] [PMC]8. Filipski E, Lévi F. Circadian disruption in experimental cancer processes. Integr Cancer Ther 2009;8:298–302.
[Article] [PubMed]9. Davidson AJ, Sellix MT, Daniel J, Yamazaki S, Menaker M, Block GD. Chronic jet-lag increases mortality in aged mice. Curr Biol 2006;16:R914–R916.
[Article] [PubMed] [PMC]10. Basu P, Tripathi R, Mishra R, Singaravel M. Effect of chronic jet lag after induction of Dalton’s lymphoma in male and female mice. Biol Rhythm Res 2012;43:331–339.
[Article]11. Bjorvatn B, Pallesen S. A practical approach to circadian rhythm sleep disorders. Sleep Med Rev 2009;13:47–60.
[Article] [PubMed]12. Kuboyama T, Tohda C, Komatsu K. Neuritic regeneration and synaptic reconstruction induced by withanolide A. Br J Pharmacol 2005;144:961–971.
[Article] [PubMed] [PMC]13. Zhu J, Park S, Jeong KH, Kim WJ. Withanolide-A treatment exerts a neuroprotective effect via inhibiting neuroinflammation in the hippocampus after pilocarpine-induced status epilepticus. Epilepsy Res 2020;165:106394.
[Article] [PubMed]14. Tomita T, Wadhwa R, Kaul SC, Kurita R, Kojima N, Onishi Y. Withanolide derivative 2,3-dihydro-3β-methoxy withaferin-A modulates the circadian clock via interaction with RAR-related orphan receptor α (RORa). J Nat Prod 2021;84:1882–1888.
[Article] [PubMed]15. Kukkemane K, Jagota A. Therapeutic effects of hydro-alcoholic leaf extract of Withania somnifera on age-induced changes in daily rhythms of Sirt1, Nrf2 and Rev-erbα in the SCN of male Wistar rats. Biogerontology 2020;21:593–607.
[Article] [PubMed]16. Ruby NF, Patton DF, Bane S, Looi D, Heller HC. Reentrainment impairs spatial working memory until both activity onset and offset reentrain. J Biol Rhythms 2015;30:408–416.
[Article] [PubMed] [PMC]17. Iggena D, Winter Y, Steiner B. Melatonin restores hippocampal neural precursor cell proliferation and prevents cognitive deficits induced by jet lag simulation in adult mice. J Pineal Res 2017;62:e12397.
[Article]18. Chen LX, He H, Qiu F. Natural withanolides: an overview. Nat Prod Rep 2011;28:705–740.
[Article] [PubMed]19. Portaluppi F, Smolensky MH, Touitou Y. Ethics and methods for biological rhythm research on animals and human beings. Chronobiol Int 2010;27:1911–1929.
[Article] [PubMed]20. Jud C, Schmutz I, Hampp G, Oster H, Albrecht U. A guideline for analyzing circadian wheel-running behavior in rodents under different lighting conditions. Biol Proced Online 2005;7:101–116.
[Article] [PubMed] [PMC]21. Pitts MW. Barnes maze procedure for spatial learning and memory in mice. Bio Protoc 2018;8:e2744.
[Article] [PubMed] [PMC]22. Cho K. Chronic ‘jet lag’ produces temporal lobe atrophy and spatial cognitive deficits. Nat Neurosci 2001;4:567–568.
[Article] [PubMed]23. Ángeles-Castellanos M, Amaya JM, Salgado-Delgado R, Buijs RM, Escobar C. Scheduled food hastens re-entrainment more than melatonin does after a 6-h phase advance of the light-dark cycle in rats. J Biol Rhythms 2011;26:324–334.
[Article] [PubMed]24. Takahashi T, Sasaki M, Itoh H, Ozone M, Yamadera W, Hayshida K, et al. Effect of 3 mg melatonin on jet lag syndrome in an 8-h eastward flight. Psychiatry Clin Neurosci 2000;54:377–378.
[Article] [PubMed]25. Pfeffer M, Rauch A, Korf HW, von Gall C. The endogenous melatonin (MT) signal facilitates reentrainment of the circadian system to light-induced phase advances by acting upon MT2 receptors. Chronobiol Int 2012;29:415–429.
[Article] [PubMed]26. Mantar F, Dilan KH. Determining the nutrition, sleep quality and body composition conditions of working shift workers in furniture production. Endocr Abstr 2020;70:AEP425.
[Article]